Sélection naturelle (GP)
Comment citer ?
Bourrat, Pierrick (2016), «Sélection naturelle (GP)», dans Maxime Kristanek (dir.), l'Encyclopédie philosophique, consulté le ..., https://encyclo-philo.fr/selection-naturelle-gp
Publié en septembre 2016
Résumé
La sélection naturelle est un mécanisme par lequel une population peut évoluer et s’adapter à son environnement. Il existe différentes façons de présenter ce mécanisme. Nous commencerons tout d’abord par le présenter en utilisant un exemple classique dans la littérature, celui de la phalène du bouleau, un papillon nocturne. Nous poursuivrons par une description plus abstraite, connue sous le nom des trois conditions de Lewontin pour l’évolution par la sélection naturelle. Ceci nous permettra de distinguer clairement le processus par lequel une population peut évoluer de l’évolution elle-même. Elle nous permettra également de mettre en évidence certaines des limitations de cette façon d’abstraire la sélection naturelle. Nous présenterons ensuite une autre conception du processus de sélection naturelle, la théorie du réplicateur que nous critiquerons brièvement. Nous discuterons enfin l’idée selon laquelle la sélection naturelle peut agir à différents niveaux d’organisation.
Introduction
La sélection naturelle est un mécanisme proposé par Charles Darwin en 1859 dans l’Origine des Espèces pour expliquer l’adaptation des organismes biologiques à leur environnement. Ce mécanisme est l’un des mécanismes pivots de la théorie moderne de l’évolution. Le mécanisme de sélection naturelle est essentiel car il est le seul mécanisme connu permettant d’expliquer l’adaptation des organismes à leur environnement sans mentionner une intervention divine. Il permet en effet d’expliquer, conjointement avec l’apparition de mutations, c’est-à-dire de nouveau variants, l’existence de structures complexes telles que l’œil. Dans une population, lorsque des mutations apparaissent, la sélection naturelle « fait le tri » en ne retenant que ceux qui sont les aptes. Ces individus les plus aptes se reproduisent et de nouvelles mutations apparaissent dans leur descendance et le cycle recommence les structures créées, de par leur complexité et la fonction qu’elles remplissent semblent parfois (comme dans le cas de l’œil) le résultat de l'intervention directe d'un créateur, alors qu’elles sont en fait le résultat d’un processus aveugle.
1. L’expérience de Kettlewells
Mais que veut dire l’expression « la sélection naturelle fait le tri » ? L’une des façons les plus simples de l’expliquer est de commencer par en donner un exemple. Supposons une population dans laquelle on trouve deux variants d’une espèce d’organismes, disons des papillons, dont l’un a une couleur différente de l’autre, disons blanc pour l’un et noir pour l’autre. Imaginons maintenant que ces papillons vivent sur des troncs d’arbres qui, pour la majorité, ont une couleur sombre et qu’il existe des prédateurs de papillons dans l’environnement qui chassent à vue. Dans cet exemple, il est assez facile d’imaginer que les papillons noirs auront un avantage face aux prédateurs par rapport aux papillons blancs : les prédateurs verront plus souvent les papillons blancs, les chasseront donc plus souvent, ce qui aura pour effet de rendre la survie des papillons blancs inférieure à celle des papillons noirs toutes choses étant égales par ailleurs. Les prédateurs représentent ici ce que l’on appelle une pression de sélection. Une pression de sélection représente le facteur qui « fait le tri » entre les variants.
Si l’on suppose maintenant que les papillons noirs et les papillons blancs ont, s’ils survivent, en moyenne le même nombre de descendants (nombre d’individus auxquels ils donnent naissance) et enfin que les deux variants transmettent leur couleur à leur progéniture, il en découle que le nombre de papillons noirs augmentera dans cette population. Il y aura donc évolution de la couleur dans la population, si bien que si l’on attend suffisamment longtemps, seuls (ou presque) les papillons noirs seront présents dans cette population. Et puisque la raison pour laquelle la couleur moyenne de la population évolue du fait que les papillons noirs ont en moyenne un avantage sur leur survie face aux papillons blancs (ils sont plus aptes), on dit que la sélection naturelle est un facteur responsable de cette évolution.
Cet exemple de sélection sur la couleur des papillons est en fait celui présenté par Kettlewells dans le milieu des années 1950 sur l’évolution de la couleur de la phalène du bouleau. Kettlewells voulait expliquer pourquoi le nombre de papillons noirs avait commencé à augmenter dans certaines régions urbaines et industrielles de l’Angleterre au cours du 19ème siècle, alors que les blancs restaient prédominants dans les régions non urbaines. Kettlewells testa l’hypothèse que la sélection naturelle était responsable de cette évolution de la couleur. Les troncs des bouleaux normalement blancs, deviennent noirs lorsque la suie (noire) s’échappant des cheminées des usines vient se déposer sur ceux-ci. Kettlewells raisonna que dans une zone polluée, la couleur noire pourrait être un avantage puisqu’elle permet un meilleur camouflage au papillon que la couleur blanche très visible. Dans une zone non polluée ce serait tout le contraire, la couleur noire sur un tronc blanc de bouleau serait un très mauvais camouflage alors que la couleur blanche permettrait un meilleur camouflage. Cette différence pourrait donc expliquer la différence de fréquence entre les deux types de papillons entre les zones polluées et non polluées. Pour tester cette hypothèse Kettlewells captura, relâcha puis captura à nouveau un certain nombre de phalènes des deux types (qui étaient marquées pour qu’on puisse les reconnaître) d’abord dans une zone polluée puis d’autres papillons, aussi des deux types, dans une zone non polluée. Il captura la seconde fois un nombre de phalènes noires supérieure aux blanches dans la zone polluée, suggérant que les phalènes noires survivaient mieux dans cette zone. Plus tard, observa le résultat contraire dans la zone non polluée, suggérant que les phalènes blanches survivaient mieux que les phalènes noires dans la zone non polluée. Ceci démontra la plausibilité de l’explication sélective.
Cet exemple met en évidence la simplicité du mécanisme par lequel la sélection naturelle peut être responsable de l’évolution et plus spécifiquement de l’adaptation dans une population. Dans le détail, le processus de sélection naturelle a fait néanmoins l’objet de questionnements théoriques importants.
2. Les conditions de Lewontin pour l’évolution par sélection naturelle
Une manière plus générale et plus précise de représenter la sélection naturelle et qui permet d’identifier certains des enjeux philosophiques de ce concept est celle proposée par Lewontin (1970) qui s’était lui-même inspiré de Darwin (1859). Lewontin propose que pour qu’une population (de papillons par exemple) évolue par sélection naturelle, il faut :
- Qu’il y ait de la variation phénotypique dans la population, c’est-à-dire que les individus aient des caractéristiques différentes. Dans le cas du papillon, la couleur est le phénotype qui varie.
- Que les différents phénotypes amènent à différents taux de survie
set/ou reproductifs dans différents environnements, ou autrement dit à des différences de fitness. C’est ce que l’on suppose dans le cas du papillon : on dira que les papillons blancs ont une fitness supérieure s’ils survivent mieux et se reproduisent plus souvent que les papillons noirs dans un environnement donné. - Que les différences phénotypiques soient héritables, c’est-à-dire que les descendants ressemblent plus à leurs parents qu’ils ne ressemblent aux autres individus. Là encore, c’est ce que l’on suppose dans le cas du papillon : les papillons blancs produisent plus souvent des papillons blancs que des papillons noirs et inversement.
Lewontin propose que si ces trois conditions sont vérifiées, une population évoluera par sélection naturelle. Il est assez facile de démontrer que cela ne sera pas toujours le cas. En fait si on peut imaginer des cas où, en dépit de variation phénotypique (première condition) amenant à des différences de fitness (deuxième condition) héritables (troisième condition), il n’y a pas d’évolution. Cela peut se comprendre si l’on se souvient que la sélection naturelle n’est qu’un des processus pouvant expliquer l’évolution d’un trait au sein d’une population. Si par exemple les différents phénotypes d’une population ne transmettent pas tous leurs traits de la même manière à leurs descendants, il se peut qu’un phénotype éliminé au cours d’une génération par la sélection naturelle réapparaisse à la génération suivante du fait d’une transmission imparfaite du phénotype.

Figure 1 : Evolution par sélection naturelle des individus noirs
Cas d’une population composée de huit individus (on peut imaginer qu’il s’agisse de papillons, mais l’exemple peut s’appliquer plus généralement) : deux blancs, quatre gris et deux noirs. On suppose que les individus gris ont une fitness inférieure à celles des individus blancs qui produisent toujours un individu chacun et que les individus blancs ont une fitness inférieure aux individus noirs qui produisent toujours trois individus chacun. Si la transmission est parfaite pour chaque type, on voit bien sur la figure qu’après une génération, il y a plus d’individus noirs (six) que d’individus blancs (deux) et gris (qui pour ces derniers ont disparu de la population).

Figure 2 : Sélection naturelle des individus noirs sans évolution
Cas identique à celui de la Figure 1, mais supposant que les individus noirs produisent en moyenne un individu noir et deux individus gris. Il y a dans ce cas de la sélection naturelle (les individus noirs ont plus de descendants que les autres) mais il n’y a pas d’évolution observée car un autre facteur contrebalance l’effet de la sélection naturelle.
Il est donc crucial de distinguer la sélection naturelle, c’est-à-dire le fait que certains individus ont un avantage qui leur permet d’avoir plus de descendants que les autres dans une population, de l’évolution, qui est le résultat d’une combinaison de facteurs incluant la sélection naturelle mais aussi d’autres facteurs, tels que la production de variation (mutation) ou d’effets aléatoires qui amènent certains individus à se reproduire plus que les autres alors qu’ils n’ont pas d’avantages particuliers par rapport à d’autres. Pour ce dernier cas, on parle alors de dérive génétique. Ce phénomène est couramment observé dans des populations de petite taille dans lesquelles des facteurs aléatoires prennent une importance prépondérante pour expliquer l’évolution des traits de ces populations. Pour fixer les idées, le phénomène de dérive est semblable à celui d’une succession de tirage à pile ou face d’une pièce non biaisée (c’est-à-dire avec une probabilité égale de tomber sur pile ou face). Si l’on tire à pile ou face quatre fois, il est possible que la pièce « par chance » tombe quatre fois sur pile, alors que l’on s’attendrait à ce qu’en moyenne elle tombe deux fois sur pile et deux fois sur face. Si l’on lance maintenant cette pièce dix mille fois, il est extrêmement peu probable que la pièce tombe dix mille fois sur pile et bien plus probable qu’elle tombe environ 5000 fois sur pile. Dans le cas de la dérive il est possible que dans une petite population où des individus d’un type qui ont une fitness identique à un autre type voient néanmoins leur fréquence augmenter « par chance » jusqu’à envahir totalement la population (Figure 3). Ces effets diminuent au fur et à mesure que la population augmente (Figure 4). Lorsqu’on fait l’hypothèse que la population est d’une très grande taille, voire infinie (ce qui est une idéalisation, ligne bleue sur les Figures 3 et 4), on peut alors considérer que la dérive génétique est nulle.

Figure 3
Simulations de l’évolution de dix populations identiques de deux variants contenant 10 individus sur 100 générations. Les deux variants ‘A’ et ‘a’ qui ont la même fitness commencent avec une fréquence de 0.5 (5 individus de chaque) à la génération 0. Le graphique représente l’évolution de la fréquence du variant A dans chaque population au cours des générations. Due à une dérive génétique forte on observe que très rapidement soit le variant A envahi la population, soit il est perdu (suivant le cas) par chance. Figure réalisée et modifiée à partir du logiciel ‘GeneP’.
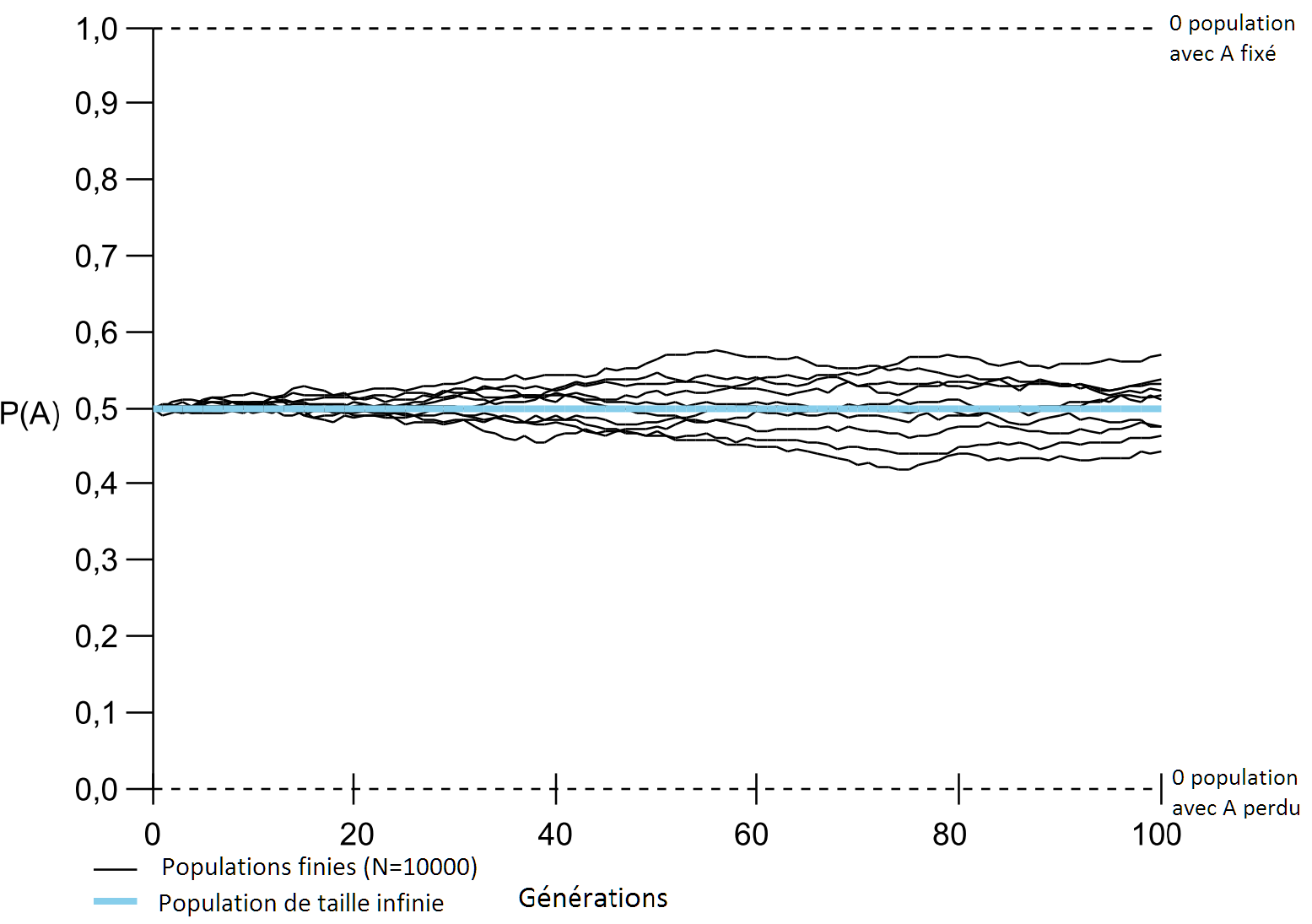
Figure 4
Simulations similaires à celles de la figure 3, mais dans ce cas la taille de chaque population est de 10000 (5000 pour chaque variant). On observe de la dérive pour chaque population, mais elle est beaucoup moins prononcée que lorsque la taille de la population est de 10 individus (Figure 3). Sur 100 générations le variant A n’a ni été perdu, ni envahi dans aucune des 10 populations. Figure réalisée et modifiée à partir du logiciel ‘GeneP’.
3. Les réplicateurs et la sélection naturelle
Les trois conditions de Lewontin sont donc une manière de représenter le processus par lequel la sélection naturelle amène à l’évolution d’une population. Il existe d’autres façons de présenter ce processus. La plus célèbre est due en grande partie à Richard Dawkins qui propose en 1976 que la sélection naturelle implique nécessairement des réplicateurs, c’est-à-dire des entités capables de se reproduire avec une très grande fidélité, ou en d’autres termes qui ne mutent que très peu. Selon cette proposition, les réplicateurs qui se reproduisent le plus vite et le plus efficacement, deviendront les plus fréquents dans une population. Ce point de vue amène Richard Dawkins à considérer que les corps des individus ne sont que des véhicules passifs qui ne servent qu’aux intérêts des réplicateurs, en l’occurrence des gènes. Dawkins suppose qu’il puisse exister des réplicateurs d’un autre type que les gènes, notamment dans le domaine culturel. Il baptise le gène culturel du nom de « mème ». Bien que l’idée de généraliser la notion de gène au domaine de la culture soit simple et élégante, elle a été vivement critiquée dans la littérature pour diverses raisons. Premièrement, personne n’a découvert le support de l’information « mémétique » analogue au support génétique qui est l’ADN, une molécule présente dans toutes les cellules des êtres vivants et qui permet à l’information génétique d’être transmise. Deuxièmement, il n’est pas clair que l’évolution culturelle résulte de la sélection plutôt que d’autres processus évolutifs. On peut en effet imaginer qu’une mélodie (un même ou plus exactement sa manifestation) n’est pas seulement répliquée comme l’ADN d’un individu à l’autre, mais qu’elle est en partie réinventée ou reconstruite par les individus. Si je siffle par exemple « au clair de la lune » avec une fausse note il est probable qu’une autre personne m’entendant siffler se mette elle-même à siffler la même mélodie mais en corrigeant la fausse note.
D’une manière plus générale, l’idée du réplicateur et plus particulièrement du gène comme unité de base de la sélection naturelle a reçu certaines critiques. L’une des plus importantes est que s’il est possible de réduire toute évolution à l’évolution d’une fréquence de gènes au cours du temps, cela ne représente pour autant pas causalement la manière dont la sélection naturelle permet l’évolution. Parler en termes de gènes de l’évolution permet de tenir les comptes, mais ne permet pas d’expliquer la raison pour laquelle ce gène est surreprésenté à la génération suivante, surtout s’il est associé à différents traits. Il est possible par exemple que les grands individus soient aussi ceux qui ont une couleur particulière, disons noire, mais que la taille ne soit pas causalement associée à la couleur. Si l’on suppose que la couleur noire ne donne aucun avantage mais que les grands individus produisent plus d’individus et que l’on fait les comptes, il y aura plus d’individus noirs. Une autre critique de cette théorie est que si la réplication est nécessaire pour la sélection naturelle, alors une origine de la sélection naturelle sans réplication semble impossible. Or il semblerait bien que la capacité à se répliquer elle-même ait évolué et qu’elle soit donc due à un type de sélection naturelle.
4. La controverse de la sélection de groupe
Nous l’avons vu les conditions de Lewontin ne supposent pas d’unité particulière sur laquelle la sélection peut agir. Le trait sur lequel agit la sélection pourrait, en effet être le phénotype d’un individu multicellulaire tel qu’un humain, une colonie d’abeille, une cellule, un chromosome, etc. Dans la théorie du réplicateur, une seule unité compte : le gène. Les autres unités d’organisation ne sont là que pour interagir avec l’environnement et permettre aux gènes de se répliquer. Bien que la théorie du réplicateur ne privilégie en principe aucune unité d’organisation au-delà du gène, ces principaux défenseurs ont souvent considéré l’individu biologique (par exemple un humain) comme étant la principale unité d’organisation biologique sur laquelle la sélection naturelle opère, en réponse à une autre hypothèse, celle proposant que la sélection naturelle opère au niveau des groupes d’individus. Selon cette dernière théorie les individus d’un groupe pourraient exprimer un phénotype qui réduirait leur succès reproducteur mais augmenterait le succès du groupe sur le long terme. Cette théorie fut largement rejetée à la suite du travail de George Williams (1966) qui démontra que les modèles sur lesquels étaient basés l’hypothèse de sélection de groupe n’étaient pas suffisamment réalistes. L’hypothèse de l’organisme individuel comme unité privilégiée de sélection n’est cependant pas sans poser problème. En effet si les individus multicellulaires ont évolué comme on le suppose par sélection naturelle à partir d’organismes unicellulaires, il semble difficile d’imaginer qu’il n’y ait pas eu une forme de sélection au niveau du groupe de cellules durant la transition entre les organismes unicellulaires et multicellulaires. Si en effet la théorie individualiste prend pour acquis l’individu multicellulaire alors qu’elle devrait en démontrer l’origine, alors cette théorie doit être repensée pour être plus générale et inclure dans son schème le fait que les individus multicellulaires sont aussi des groupes de cellules.
Bibliographie
Darwin, C. 1859. L’Origine Des Espèces Au Moyen de La Sélection Naturelle: Ou La Préservation Des Races Favorisées Dans La Lutte Pour La Vie. Editions Flammarion.Livre dans lequel on trouvera les idées originales de Darwin sur la sélection naturelle.
Dawkins, R. 1976. Le Gène Égoïste. Paris: Odile Jacob.Présentation moderne de l’idée de sélection naturelle depuis le point de vue du gène. Probablement le livre qui a eu le plus d’influence sur la biologie évolutive moderne depuis les 40 ans dernières années.
Kettlewell, H.B.D. 1955. “Selection Experiments on Industrial Melanism in the Lepidoptera.” Heredity 9 (3): 323–42.En anglais. Compte rendu scientifique de la première expériences sur la phalène du bouleau de Kettlewell dans une zone polluée.
Lewontin, R. C. 1970. “The Units of Selection.” Annual Review of Ecology and Systematics 1: 1–18.En anglais. L’article original de Lewontin ou l’on trouve la formulation des trois conditions pour l’évolution par sélection naturelle.
Williams, G. C. 1966 Adaptation and Natural Selection. Princeton: Princeton University Press.En anglais. L’ouvrage classique de George Williams.
Pour aller plus loin
Huneman, Philippe. 2009. “Sélection.” In Les Mondes Darwiniens, 47–86. paris: Syllepse.
PopG version 4.02 développé par Joseph Felsenstein
Lien pour télécharger le logiciel avec lequel les figures 3 et 4 ont été réalisées : http://evolution.gs.washington.edu/popgen/popg.html.On peut l’utiliser pour simuler l’évolution de populations avec deux variants sous l’influence entre autres de sélection et dérive.
Pierrick Bourrat
The University of Sydney